带有分化潜能的高扩增能力:即使在传代多达10次后,MSC仍然能够保持其增殖和多向分化能力。然而,负责这种特性的干细胞亚群还未知。
可以通过释放可溶性因子来调节免疫能力
许多研究表明,MSC可能是具有多种功能的异质细胞混合物。目前体外也发现了源自MSC的三能、双能和单能克隆,意味着MSC在克隆形成和多向分化方面存在显著的异质性。
BMMSCs和WJMSCs的单细胞分群作者使用了第6-7代体外扩增的BMMSC和WJMSC,因为这些细胞可以保持在稳定状态并已经用于临床。拿到不同供体的3个WJMSC和3个BMMSC样本进行了10X的scRNA测序,获得了61296个MSCs(33594个WJMSCs和27702个BMMSCs)。
降维聚类得到了6群细胞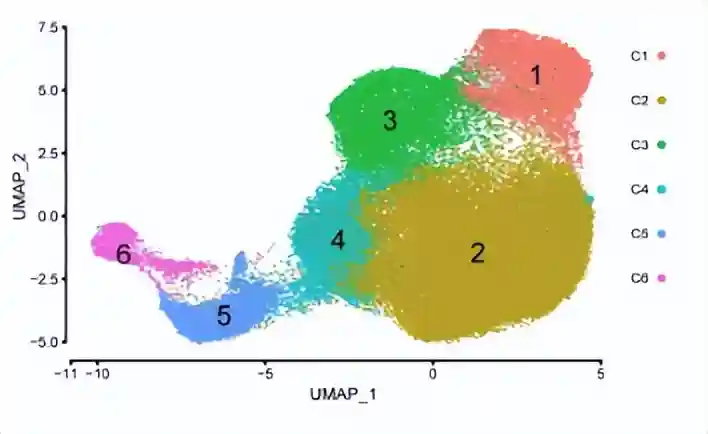
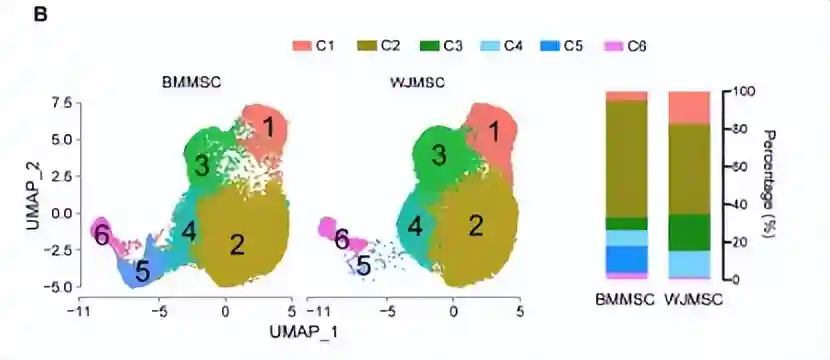
发现WJMSC相比于BMMSC,具有更高的供体间异质性
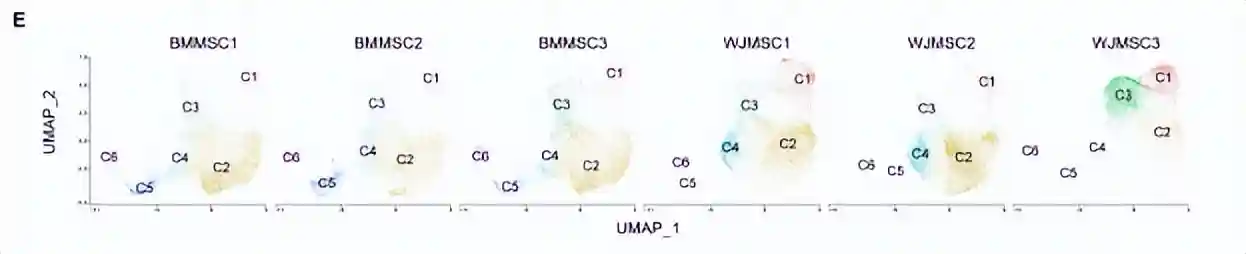
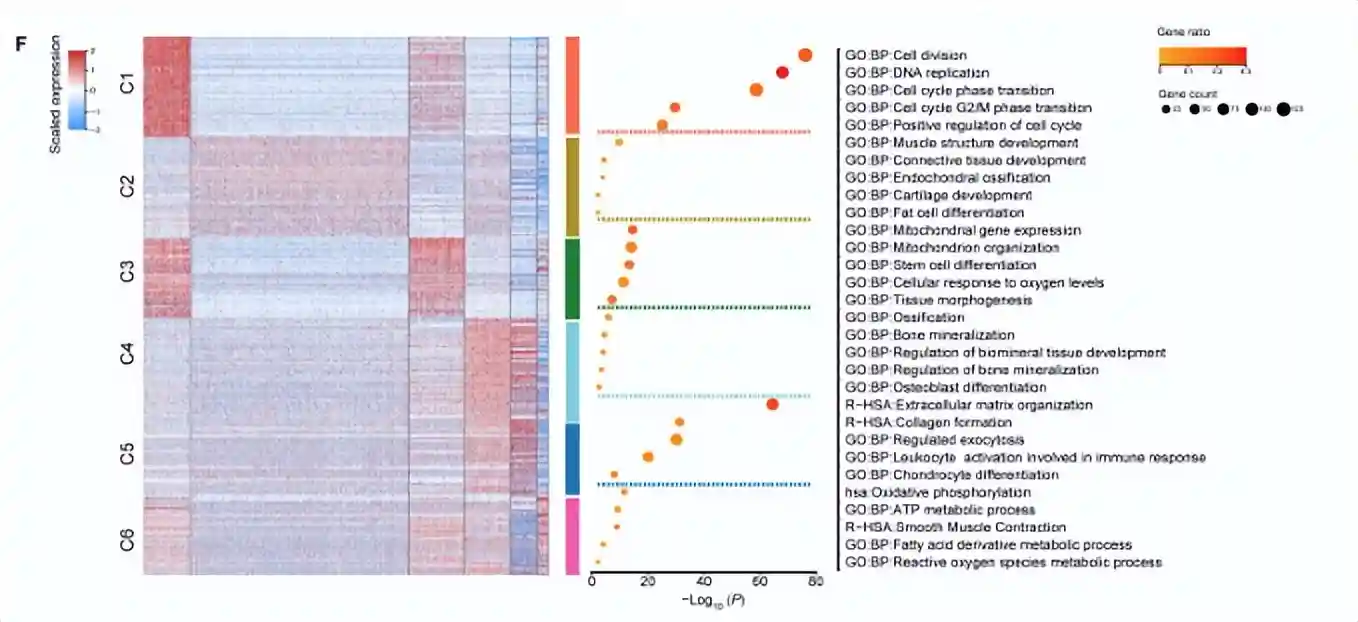
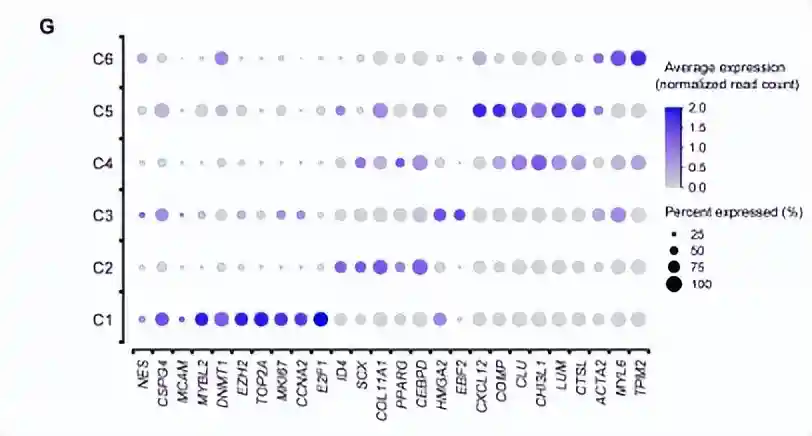
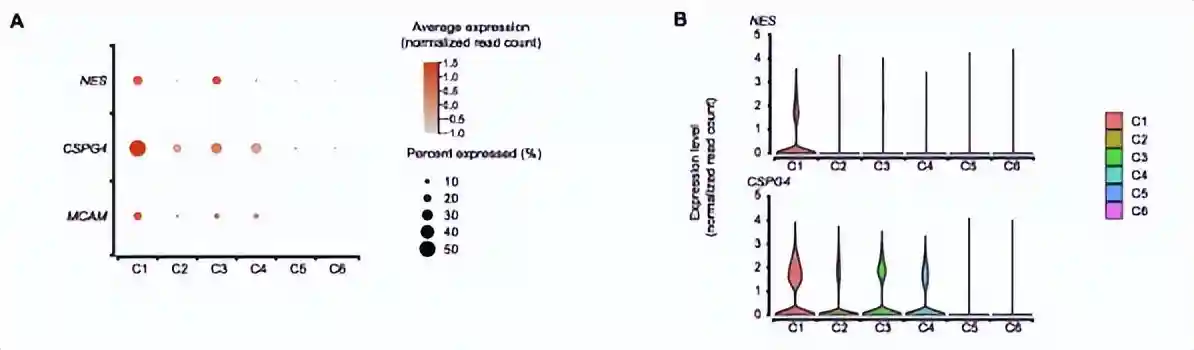
cluster1高表达与DNA复制和细胞周期相关的基因,包括增殖(TOP2A、MKI67)和E2F1)和细胞周期调节(CCNA2)的marker基因。有趣的是,NG2/CSPG4、CD146/MCAM和NES是血管周围中胚层祖细胞的marker基因,也在cluster1的细胞中高表达。此外,维持多能性和未分化干细胞状态所必需的基因(MYBL2、DNMT1和EZH2)的高表达,从侧面反映了cluster1可能是stem-likeAPC。
cluster2占据总细胞数的一半以上,高表达分化相关的基因。例如成骨分化(ID4)、软骨分化(SCX和COL11A1)和细胞成脂分化(PPARG和CEBPD)。它具有多向分化潜能,于是被称为tripotentmultipotentMPCs。
除了干细胞和祖细胞外,还发现了分化的前体细胞。cluster3的细胞富含参与干细胞分化(PSMD2、PSMD7和PHF5A)、组织形态发育(TBX3、CFL1和TRIM28)和脂肪形成相关的线粒体发育(NDUFA9、UQCC2、ATP5F1B和COX20)的基因。此外,Cluster3细胞高表达脂肪细胞分化相关的基因,例如EBF2和HMGA2。因此,cluster3被称为单能前脂肪细胞。
cluster4中的细胞高表达软骨特异性基因COMP和细胞外基质重塑基因CHI3L1、CLU、LUM和CTSL;另外还发现了成骨相关基因的高表达(OMD、ASPN、GPM6B、IFITM1和GPNMB),因此与软骨细胞和成骨细胞密切相关。

cluster5类似于软骨细胞,并且高表达软骨形成相关基因(COL6A3、COL6A1和ECM1);有趣的是,它的高表达基因还涉及了免疫调节和分泌的通路。因此它被定义为免疫调节前软骨细胞。
cluster6的细胞,占细胞总数的比例最低(2.25%),高表达平滑肌收缩所必需的基因(ACTA2、MYL6和TPM2)。因此称为前平滑肌细胞(pre-SMC),这与MSC向血管谱系分化的结果一致。
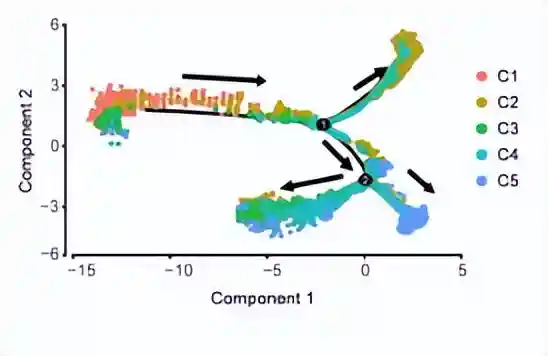
为了探索细胞群之间的关系和发育次序,作者使用Monocle2进行了拟时序推断。
发现首先是stem-likeAPCs处于最早的细胞位置,然后是MPC。之后MPC衍生了两条分支:一条通往prechondro-osteoblasts(pre-COs)和分化的单能前软骨细胞,另一条通往前脂肪细胞。这个结果与RNA速度分析结果一致。
因此推断:细胞群从stem-likeAPCs发展为三能MPC,并最终发展为双能和单能前体。
相比于其他的clusters,Cluster1细胞高表达了NG2/CSPG4、CD146/MCAM和NES。此前报道过NESTINbrightNG2/CSPG4+periarteriolarmesodermalprogenitorcell可以进行自我更新和分化为多个间充质细胞谱系,可以分化为MSCs并构成多个器官MSCs的起源。
作者又分别比较了cluster1与NG2+小动脉周细胞、LEPR+窦周细胞和PXM细胞之间的单细胞转录组数据。利用参与维持细胞干性的基因进行相关性分析,这些基因包括了细胞自我更新基因(E2F8、CTCF、PBX3和MYBL2)、细胞分化负调节基因(ASPM、CBFB、SUZ12、WNT5A和PTHLH)和细胞周期调节基因(CDKN2C和CDKN1A)。相比于其他的clusters,cluster1与NG2+小动脉周围细胞(体内)和PXM细胞(体外)密切相关;并且细胞干性基因及notch信号通路相关基因(E2F1、EZH2、TFDP1)、细胞凋亡负调控基因(HMGB2、BRCA1、PAK4、MAZ)都在cluster1中高表达。
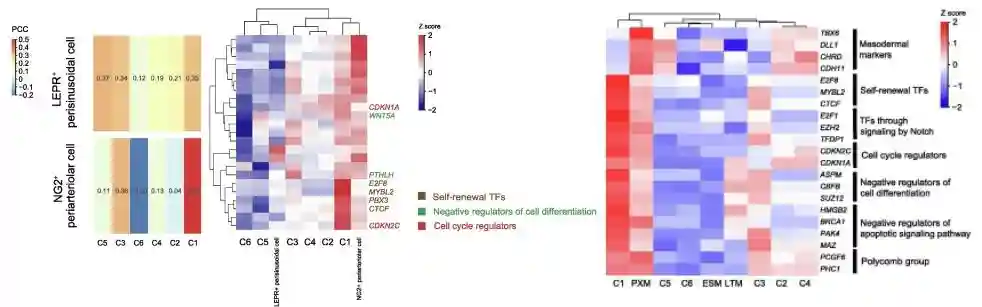
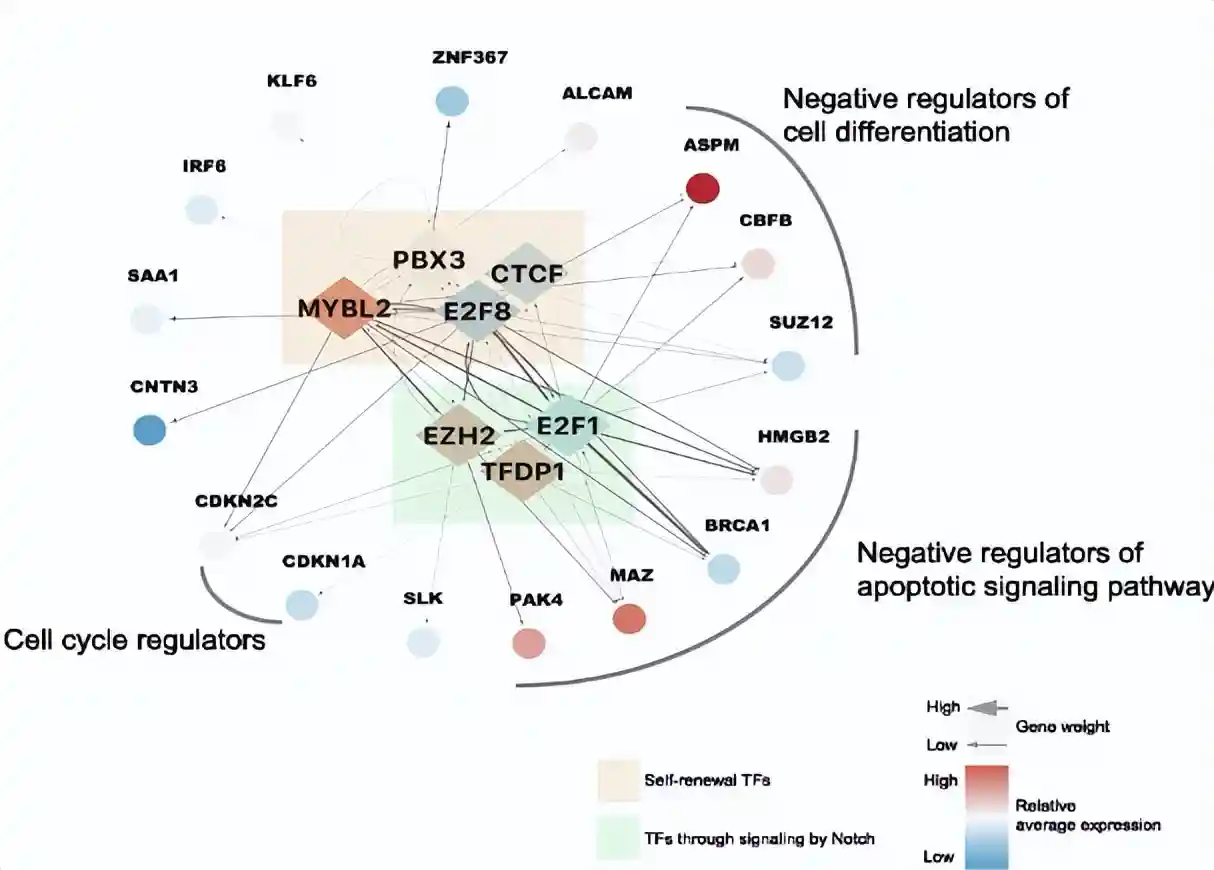
SCENIC分析发现高表达的CTCF、EZH2、E2F8、PBX3、MYBL2和TFDP1也是cluster1的转录因子编码基因,这些转录因子可以激活自我更新的生物学途径。
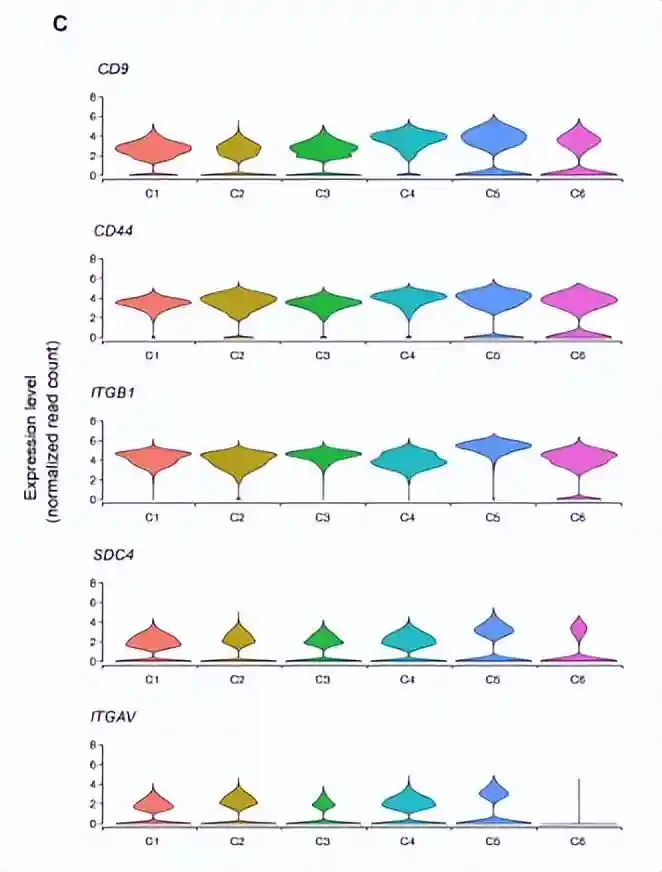
之前cluster2定义为多能MPC,并且表达了MSC细胞干性相关的基因(CD9、CD44、ITGB1、SDC4和ITGAV)
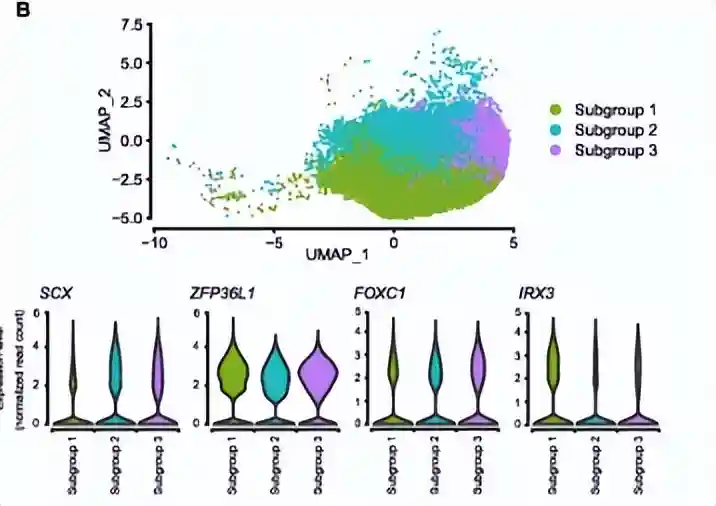
为了探索MPC的多能性是单个细胞群还是不同细胞群导致的,作者对cluster2继续细分得到3个亚群,发现除了参与中胚层发育的基因(ZFP36L1、FOXC1、IRX3和SCX)均表达外,还有其他几类特异性表达模式:
亚群1中的细胞表达与成骨细胞分化(IFITM1和TMEM119)和成骨细胞祖细胞增殖(COL1A1、ID3和ID4)相关的基因,因此称为osteo-primedMPCs
亚群2中的细胞表达与脂肪酸代谢和脂质积累相关的基因(IGF1、PPARG、FABP3、P2RY6、PTGER2和PTGFR),被称为adipo-primedMPCs
亚群3中的细胞表达软骨形成的关键基因,包括软骨细胞分化、软骨发育和胶原纤维组织(COL11A1、COL12A1、MAF、NFIB、TGFBR2、TRPS1、FGF18和INSR),被称为chondro-primedMPCs
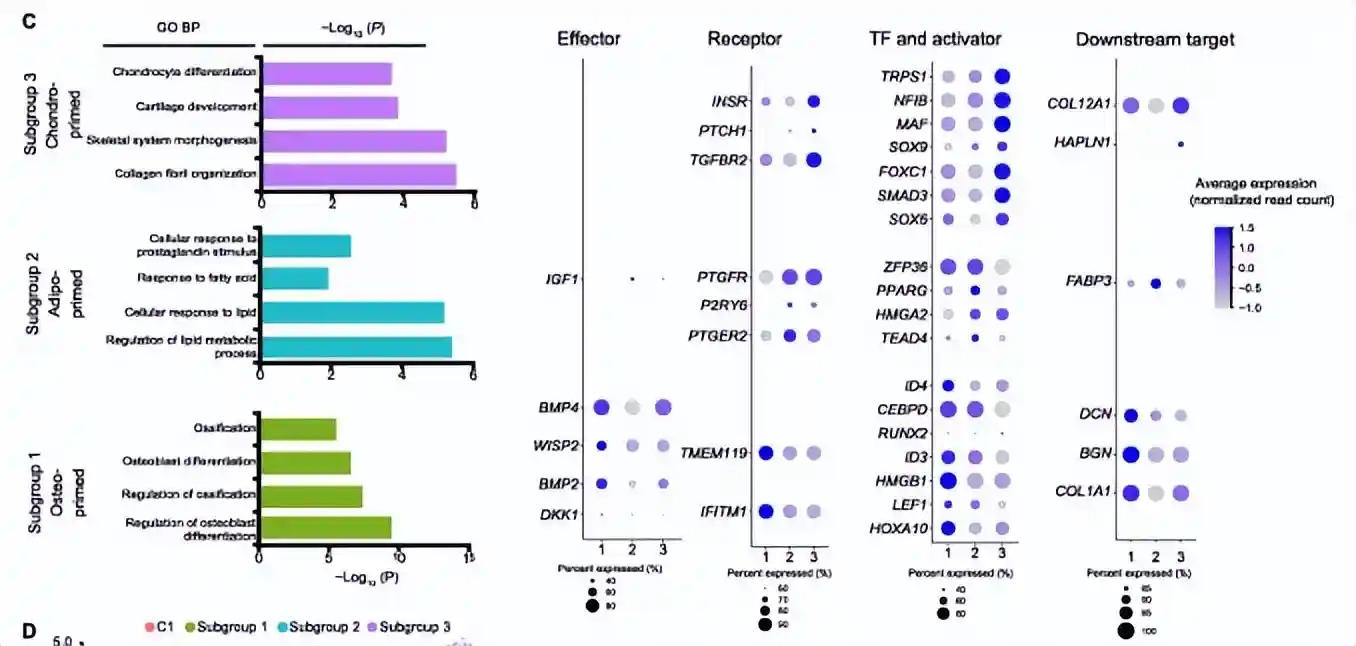
之后对这三个亚群进行轨迹分析,揭示了从最初的stem-likeAPC到MPC的三个亚群的两条主要分化途径,并且每条途径都与一个以上的MPC亚群相关。又发现了4种基因表达的模式:
第1种模式与细胞干性维持相关,并在prebranchedAPCs中高表达,其中包括细胞干性相关分子(CBFB和PTN)和编码蛋白酶体的基因(PSMB9、PSMC3、PSMD2、PSMD7和PSME2),它们在调节干细胞的自我更新、多能性和分化中发挥关键作用
模式2和模式3富含软骨特异性基因,例如SOX6、SOX9和COL12A1,以及骨特异性基因ATF4和RUNX2
模式4包含编码醛酮还原酶(AKR1C1和AKR1C2)和EPHX1的基因,它们对脂肪细胞分化至关重要
因此发现MPC存在于干细胞活性下降和持续发育为成骨细胞/软骨细胞/脂肪细胞之间的过渡状态。
参与软骨形成过程(包括硫酸软骨素分解代谢过程、软骨内骨形态发生和骨骼系统发育)的基因在cluster5中高表达。

另外,cluster5参与促炎特征的过程,包括免疫原性、补体系统激活或抑制、髓样白细胞激活,以及抗炎特征,如抑制增殖、分化和激活免疫细胞(例如,T细胞、B细胞、NK细胞和树突状细胞)
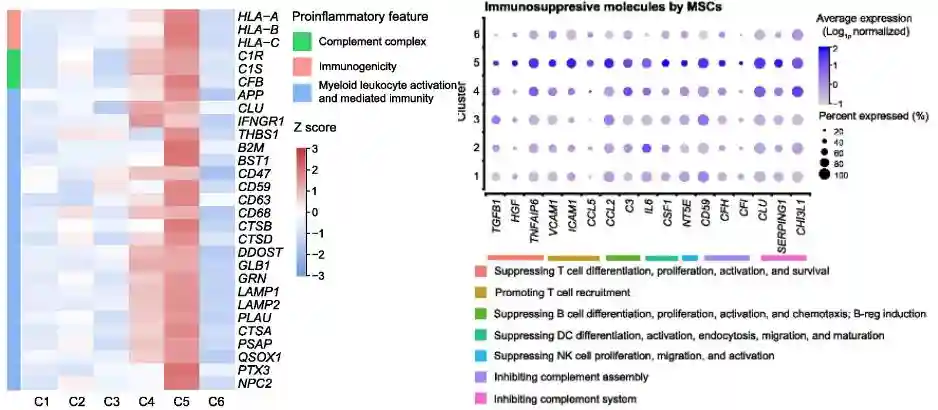
预测的转录因子包括了IRF1、NFATC2和NFKB2,这些都是先天性和获得性免疫反应的重要调节因子。
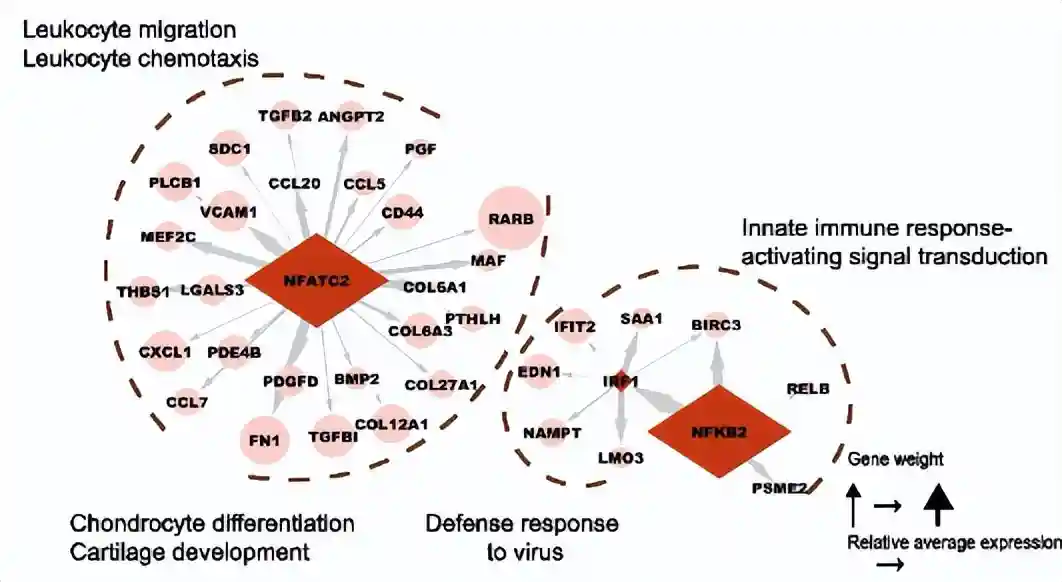
为了进一步阐明cluster5细胞如何调节炎症和免疫反应,作者使用g:Profiler进行了富集分析。结果表明,大多数免疫调节基因位于细胞外空间或细胞外囊泡中。
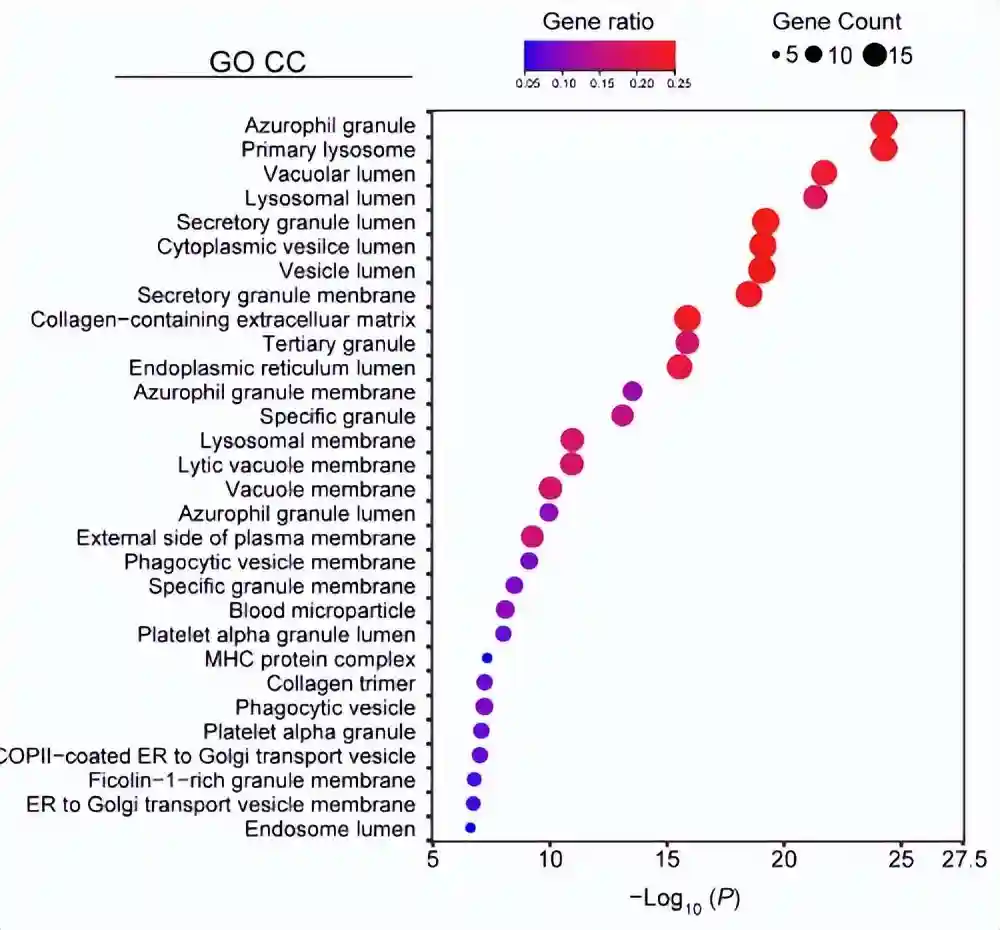
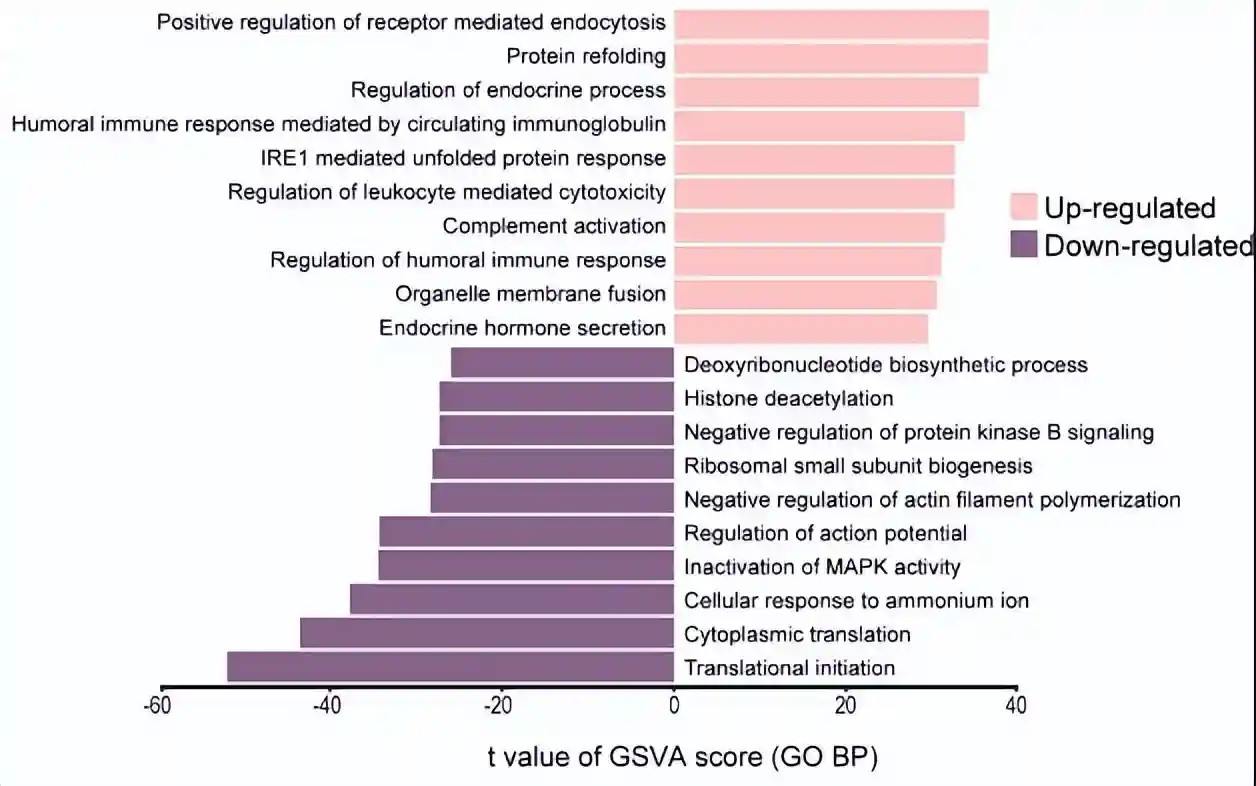
GSVA揭示了主要富集的通路,例如受体介导的内吞作用的正调节、内分泌过程的调节、细胞器膜融合和内分泌激素的分泌。
总之,这些结果表明,cluster5细胞的免疫调节作用可能是由于它们产生外泌体或分泌可溶性因子。
由于自我更新能力、多向分化潜能、旁分泌作用和免疫抑制特性,MSC被认为是基于细胞的再生医学的有前途的候选者。然而,MSC群体包含了哪种不同功能的异质性亚群仍然很大程度上未知。在这项研究中,作者对BMMSCs和WJMSCs进行了单细胞转录组分析,确定了MSCs的细胞组成,包括stem-likeAPC亚群、多能祖细胞亚群、脂肪细胞前体亚群、骨软骨细胞前体亚群和免疫调节前软骨细胞亚群,确定了特定的免疫调节亚群,不过还要在未来详细探索其免疫调节作用的机制。